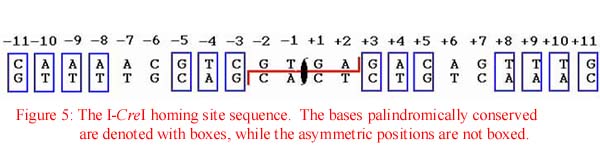
In vitro binding assays conducted by various laboratories have demonstrated that I-CreI binds to a specific homing site sequence that extends across 19-24 base pairs. The native homing site, as found in the C. reinhardtii rRNA gene, displays limited, twofold sequence symmetry around four central base pairs that are asymmetrically cleaved to generate four base, 3'OH-extended cohesive ends. Although I-CreI naturally cleaves this degenerate palindromic homing site, as shown in Figure 5, the homodimeric enzyme can bind and cleave true palindromes derived from this sequence with equal efficiency. As expected in an enzyme homodimer, the most specific points of nucleotide contact correspond to the palindromic position within the homing site.
Previous experiments have also shown that the enzyme is capable of tolerating a single base pair change at any position in the homing site. Additionally, the protein may still cleave DNA with several mutations if the mutations are of a specific identity.
A closer analysis of the cleavage site has allowed researchers to deduce that protein makes a total of 14 direct and indirect hydrogen-bond contacts to nucleotide bases on each half-site. As expected, these amino acids capable of hydrogen bonding with the DNA are donated by the beta strands of the enzyme. Furthermore, a total of 12 phosphate groups from the substrate are in contact with other residues of I-CreI. Base pairs that do not exhibit direct protein contacts are highly insensitive to various mutations -- thus confirming previous predictions that the identity of contact sites are imperative to the function of the protein.
I-CreI must attribute most of its DNA cleaving abilities to the all-important LAGLIDADG motif. First of all, this motif plays a critical role in protein folding by pointing the hydrophobic residues of the helices directly into the protein core. This helps in dimerization and domain packing -- ultimately stabilizing the structure of the enzyme. But more importantly, this motif is intricately involved in I-CreI's strategy of cleavage. Since this enzyme cleaves across the minor groove of the DNA substrate, I-CreI must deal with the steric hindrance that results from phosphate groups of an undistorted DNA double helix, which are about 10 angstroms apart. Any phosphodiester bond cleavage event, as depicted in Figure 6, must require the following:

1) An attacking nucleophile.
2) An electrophile (the phosphorus atom).
3) An leaving group (a hydroxyl oxygen).
4) A proton donor for the leaving group.
5) A group to activate the nucleophile (usually a metal or metal-bound water).
6) A group to stabilize the developing negative charge of the transition state.
By using glycines as amino acid residues, the LAGLIDADG motif allows the enzyme to position all the necessary catalytic machinery around each phosphate group in the constrained minor groove.
Studies of I-CreI bound to it homing site in the presence of inhibitory calcium ions have lead scientist to propose a possible mechanism for hydrolysis whereby a single metal ion activates a water for nucleophilic attack and stabilizes the pentacoordinate phophoanion transition state through direct interaction. This mechanism follows the schematic shown in Figure 6. From Figure 7 below, we see an image of the active site of I-CreI bound to its DNA substrate.
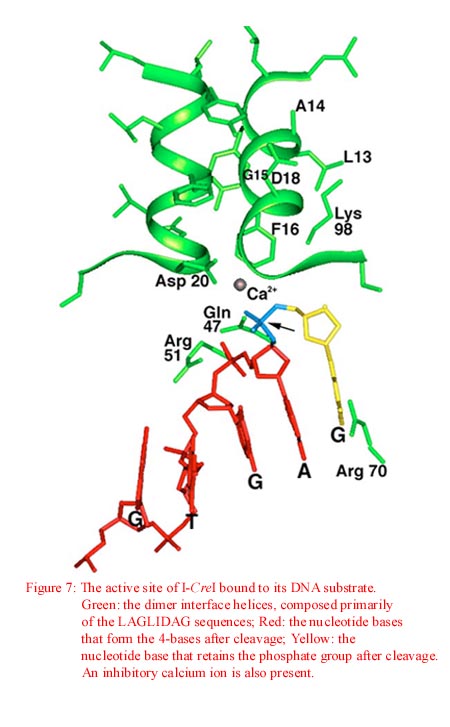
As can be seen, the carboxylate oxygen of the Asp 20 side chain (donated by the LAGLIDADG motif) coordinates with the metal ion. The oxygen of the labile phosphate and the oxygen directly across the minor groove from the cleavage site also coordinate the metal ion. A water molecule (not shown), is bound to the metal and interacts with the amide oxygen of the Gln 47 side chain. The residues believed to stabilize the phosphate leaving group are Lys 98 and 98'. We also see that the catalytically essential Arg 51 interacts with the phosphate group immediately 5' of the phosphate group about to be cleaved while Arg 70 contacts the nucleotide base immediately 3' of the same phosphate group. As previously mentioned, the cleavage event results in DNA with 3' overhangs.