Mechanism
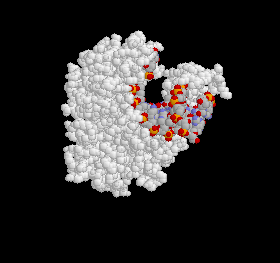
Figure 1: The Polymerase Active Site. The protein is pictured in white and the DNA strand is variously colored.
The general form of DNA is consistent throughout most organisms, so it should come as no surprise that all polymerases a certain degree of similarity. Two residues, Asp705 and Asp852, in fact, are universally conserved (7). All polymerases contain the same group of important side chains, which always include these two carboxylates (5).
Changing certain side-chains often affects certain reaction constants and these implicate the residue's involvement in replication. For instance, if changing a residue affects the Km, then we expect that it will be part of the palm where the DNA molecule binds to the polymerase. On the other hand, if the change affects kcat, this would suggest that the residue takes part in the actual polymerase reaction. Studies have demonstrated that Tyrosine, Arginine and Asparagine residues in the active site have significant interactions with the incoming molecule, while different Arginine, Glutamine and Asparagine side-chains are involved in the polymerase activity (7). These interactions are depicted in the following figures. Please note that all crystal structures are taken from Bacillus stearothermophilus and not E. coli. The corresponding residue numbers for the Klenow fragment are in parentheses:
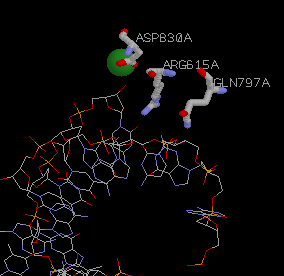
Figure 2: Arg 615(668), Gln 797(849), and Asp 830(882) provide steric complementarity to the active site. They are the most important residues for catalysis and have the greatest effect on kcat. The green cirlce is Mg2+. It's interactions with Asp830(882) are crucial to the mechanism. See Figure 4.
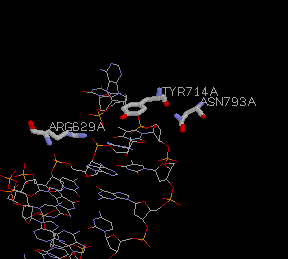
Figure 3: These residues interact more generally with the sugar backbone of the DNA structure. Tyr 714(766), Arg 629(682), and Asn 793(854) are vital to substrate binding and have great influence over the Km.
Since the active site binds A-form DNA, the B-form DNA must undergo a conformational switch upon entering the catalytic cleft. The duplex DNA must make a sharp turn as it enters the catalytic cleft. The fingers of the site bind and orient the template strand. For this interaction, the carboxylates are crucial for the polymerase reaction which is essentially a phosphoryl transfer reaction involving nucleophillic attack by the 3'-OH of the primer terminus on the dNTP a-phosphate, with release of PPi (5).
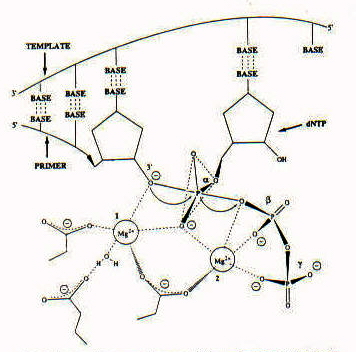
Figure 4: This is the intermediate in the mechanism proposed for the polymerase reaction. Catalysis is mediated by two divalent metal ions. You might notice the importance of Asparagine in catalysis. It interacts with each of the metal ions which, in turn, interact with the phosphate groups on the template strand.
The magnesium ligands provide substantial transition state stabilization of the negative charge on the phosphate oxygens of the pentacovalent intermediate (7). As you can see in the diagram, the alpha-phosphate is the site for nucleophilic attack. This is consistent with the expectations derived from the kinetic experiments that Asp882 would be involved with the catalytic center (5).
All of the crystal structures that I have used in Section IV have been from Bacillus stearothermophilus. The B. stearothermophilus enzyme is relatively similar to the E. coli DNA Polymerase I, although the Bacillus fragment is more processive; it incorporates 112 nucleotides compared with 7.7 nucleotides for the Klenow fragment (6). The study done by Kiefer, et al. obtained results consistent with those using other polymerase enzymes. They found that polymerase activity involves "extensive interactions with the first four to five base pairs in the minor groove, location of the terminal base pair in a pocket of excellent steric complementarity favouring correct base-pair formation, and a conformational switch from B-form to underwound A-form DNA at the polymerase active site" (5).